L’idée que l’être humain a des traces dans son ADN de néenderthalien commence à se généraliser, mais en fait l’hybridation est encore plus complexe et on commence à peine à entrevoir l’influence que cela peut avoir sur nous, sur notre résistance à certaines maladies ou au contraire, mais il semble que cela aille jusqu’à des traits de comportement. Encore un domaine où l’essor de nos connaissances bouleverse totalement notre conception de l’humanité, de sa relation à l’animalité, à la nature, à notre propre nature et apporte des potentialités nouvelles à peine entrevues avec Darwin mais je crains que ce scientifique qui a déjà essayé d’expliquer aux colonialistes de Grande Bretagne que leur ancètre local était noir, qu’il se fasse d’illusion sur la capacité à l’impérialisme occidental à ne pas tenter au contraire d’introduire des différences en matière d’hybridation. (note et traduction de danielle Bleitrach pour histoireetsociete)
Aguiche: Entretien avec le professeur Chris Stringer, l’un des plus grands experts de l’évolution humaine. ParJan Ritch-Frel Bio de l’auteur:Jan Ritch-Frel est le directeur exécutif de l’Independent Media Institute. Source: Institut indépendant des médias Crédit :Cet article a été produit par Human Bridges, un projet de l’Independent Media Institute. Tags:Afrique, Afrique/Afrique du Sud, Droits des animaux, Art, Asie, Asie/Indonésie, Asie/Philippines, Education, Europe/France, Europe/Géorgie, Europe/Espagne, Europe/Royaume-Uni, Soins de santé, Histoire, Droits de l’homme, Politique identitaire, Interview, Médias, Amérique du Nord/États-Unis d’Amérique, Océanie, Océanie/Australie, Océanie/Papouasie-Nouvelle-Guinée, Opinion, Philosophie, Science, Sciences sociales
Il y a un changement de paradigme en cours dans notre compréhension des 4 derniers millions d’années de l’évolution humaine : notre histoire est une histoire qui inclut des combinaisons avec d’autres espèces d’Homo, réparties de manière inégale dans les populations d’aujourd’hui – pas une progression évolutive nette et linéaire.
Les progrès technologiques et un nombre croissant de preuves archéologiques ont permis aux experts dans l’étude des origines humaines et de la préhistoire d’offrir un aperçu de plus en plus clair, bien que complexe, du processus biohistorique qui a produit la population humaine et les cultures d’aujourd’hui.
la plupart du temps, de nouvelles découvertes sont présentées publiquement comme des éléments de nouveauté intéressants dans les nouvelles et la vulgarisation scientifique. L’image plus complète, et l’idée que cette information a des implications précieuses pour la société et pour nos conceptions politiques, n’atteint généralement pas la conscience publique, ou les centres d’influence.
Mais il y a une prise de conscience émergente dans la communauté des experts que l’humanité peut grandement bénéficier si elle utilisait cette connaissance comme un pilier de l’éducation humaine – et s’habituer progressivement à une compréhension fondée sur des preuves de notre histoire, de notre comportement, de notre biologie et de nos capacités. Tout indique qu’une meilleure compréhension de nous-mêmes renforcerait l’humanité dans son ensemble et rendrait la connexion et la coopération plus faciles.
Le processus prendra des décennies pour s’enraciner, et il semble que la meilleure façon à ce stade pour accélérer ce processus est de donner une vue d’ensemble de la situation et de donner aux gens des points d’ancrage clés et des points de référence scientifiques pour la compréhension.
J’ai voulu le faire en discutant de certaines des conclusions parmi les plus importantes qui émergent de la recherche avec le professeur Chris Stringer, qui a été à l’avant-garde de la compréhension de l’évolution humaine depuis des décennies. Stringer a aidé à formuler le modèle « Out of Africa » des origines de notre espèce et continue de poursuivre des projets pionniers au Musée d’histoire naturelle du Royaume-Uni à Londres en tant que chef de file de la recherche sur les origines humaines au Département des sciences de la Terre.
Jan Ritch-Frel: Un bon point de départ est que nous savons que les humains d’aujourd’hui sont le fruit d’une progéniture fertile avec des espèces Homo relatives qui s’étaient séparées de nous il y a des centaines de milliers d’années, et cela s’est poursuivi avec des espèces ancêtres aussi loin que les scientifiques sont en mesure de retracer. Ceci crée un contexte qui veut que pour les espèces de primates, il ait été possible de produire une progéniture fertile avec d’autres espèces en partageant un ancêtre commun remontant à 2 millions d’années – avec une probabilité généralement décroissante de succès au fil du temps et de divergence entre les espèces d’Homo.
Chris Stringer : Nous savons que notre espèce a produit une progéniture fertile avec les Néandertaliens et les Denisovans. Nous avons également des preuves négatives qu’il y avait des limites à l’infertilité entre certaines espèces d’Homo parce que nous n’en trouvons pas beaucoup plus de preuves dans nos génomes (du moins au niveau auquel nous pouvons le détecter) – donc les accouplements entre des espèces plus éloignées n’ont pas eu lieu, n’étaient pas fertiles, ou nous ne pouvons pas les détecter au niveau de notre technologie actuelle.
Il y a des barrières, et nous savons que dans nos génomes aujourd’hui, il y a des zones de déserts où il n’y a aucun ADN néandertalien et dénisovien. Et nous savons que certains de ces déserts se trouvent dans des zones qui influencent des caractères comme la parole et la vocalisation, et le fonctionnement du cerveau. Il est également suggéré que les enfants de sexe masculin pourraient avoir été moins fertiles ou infertiles que les filles de ces accouplements hybrides. Au niveau où nous pouvons le détecter, il n’y a pas de preuve solide jusqu’à présent d’infertilité entre Homo sapiens et nos parents plus éloignés tels que Homo floresiensis ou Homo naledi.
Nous ne connaissons donc pas encore toutes les espèces d’Homo qui auraient pu s’hybrider ou s’hybrider au cours des 2 derniers millions d’années, mais certaines d’entre elles auraient certainement été interfertiles. Nous savons que nous, les Néandertaliens et les Denisovans, étions interfertiles, par exemple.
Ritch-Frel: A partir de ce que vous dites là, les coordonnées sur la manière dont nous expliquons l’évolution humaine à nous-mêmes change– pas une progression linéaire, mais une série de combinaisons, de différents groupes qui contiennent parfois des avantages pour la survie. Dans certains cas, la survie d’une population migratrice d’Homo pourrait être facilitée par l’hybridation avec une espèce résidente qui avait survécu dans une région pendant des centaines de milliers d’années ou plus, en prenant en charge ses adaptations – au système immunitaire, à la capacité de traiter l’oxygène ou à d’autres traits – sans parler de l’échange d’informations sur la culture et le mode de vie.
Plus on en apprend à ce sujet, plus il est facile de voir que le temps est mieux considéré en tant que simple ingrédient de l’histoire de l’évolution humaine. Dans cet esprit, il est plus facile de saisir jusqu’où le concept de « primitif » peut nous amener à nous comprendre nous-mêmes et notre processus évolutif.
Alors que le monde commence à intégrer cette information au centre de l’éducation humaine, il est si important de trouver les mots racines du mieux auquel nous pouvons aspirer.
Stringer: « Archaïque » et « moderne », « humain » et « non-humain » – ce sont tous des termes chargés. Qu’est-ce qu’un humain ? Et il existe de nombreuses définitions différentes de ce qu’est une espèce.
Il y a des gens qui n’utilisent « humain » que pour les sapiens, et alors les Néandertaliens ne seraient même pas humains. Je ne suis pas d’accord avec cela, car cela signifie que nous nous sommes accouplés avec des « non-humains » au cours des 50 000 dernières années, ce qui, je pense, rend la conversation très difficile.
À mon avis, le terme « humain » équivaut à être un membre du genre Homo. Je considère donc les Néandertaliens, les rhodésiensis et les erectus comme étant tous humains.
Et les termes « moderne » et « archaïque » – ce sont des termes mal appropriés. Et j’ai essayé de m’en éloigner maintenant parce que d’une part, le terme « moderne » est utilisé pour le comportement moderne, et il est également utilisé pour l’anatomie moderne, donc ces termes entretiennent la confusion. Par exemple, certaines découvertes de fossiles humains anciens ont été décrites comme « anatomiquement modernes » mais pas « comportementalement modernes » – je pense que c’est trop déroutant pour être utile.
Quand nous regardons les premiers membres d’une espèce Homo, au lieu d’avoir le terme « archaïque », comme dans avoir des « traits archaïques », je pense que c’est plus clair si nous utilisons le terme « basal ». Basal nous met sur un chemin sans la confusion et le bagage qui peuvent venir avec des termes comme « archaïque », « primitif » et « moderne ». Dans cet usage, « basal » est un terme relatif, mais au moins un où nous pouvons trouver des critères (tels que les traits squelettiques) pour le délimiter.
Il est utile ici de considérer le processus évolutif en dehors de l’Homo sapiens. Les Néandertaliens ont également eu un processus d’évolution à partir de la période où ils se sont séparés de notre ancêtre commun. Les Néandertaliens à la fin de leur époque étaient très dérivés, très différents de la façon dont ils ont commencé il y a potentiellement 600 000 ans, et pourtant, selon la pensée conventionnelle, ils sont appelés « archaïques » (par rapport à nous, les « modernes »). Au cours de centaines de milliers d’années, ils ont développé un certain nombre de nouvelles caractéristiques physiques qui n’existaient pas dans l’ancêtre commun avec Homo sapiens. Par exemple, ils ont développé un visage qui a été tiré vers l’avant au milieu, une forme crânienne sphérique en vue arrière – même certains des os de l’oreille avaient une forme différente. Et comme nous, ils ont développé un cerveau plus gros. Les Homo neanderthalensis dérivés étaient très différents de leurs ancêtres 300 000 ans plus tôt.
Abandonnons donc le cadre verbal de « primitif » et « archaïque » et « moderne » et optons pour « basal » et « dérivé » le long de notre lignée et de celle de Néandertal.
Ritch-Frel: Un autre changement récent dans la compréhension est l’histoire de la façon dont nous avons appris à marcher. Un nombre croissant de recherches suggère que cela s’est produit sur les branches des arbres et que nos bras avaient un rôle à jouer dans l’équilibre.
Stringer: Quand vous regardez les orangs-outans et les gibbons, qui sont nos proches parents vivants en Asie du Sud-Est, nous voyons que lorsqu’ils sont dans les arbres, ils marchent déjà debout, et ils se ramifient. Certaines des feuilles et des fruits les plus tendres sont à l’extrémité des branches, donc en utilisant leurs bras plus longs, ils marcheront réellement le long des branches, se soutenant en tenant avec une ou deux mains à la branche au-dessus. Et puis ils peuvent aussi sauter facilement des extrémités des branches à l’arbre suivant, pour continuer à se nourrir.
Le point de vue est donc qu’il s’agit d’un physique pré-adapté à la bipédie. Leur corps est déjà partiellement adapté à une posture verticale et le bassin est déjà dans une situation où ils peuvent se soutenir sur deux jambes. L’idée de travail serait que nos ancêtres sont passés par une étape similaire où ils marchaient dans les branches, se nourrissaient dans les arbres, commençant régulièrement à mettre leur corps en position verticale. Et puis quand ils descendent entre les arbres, les arbres peuvent commencer à s’éclaircir si les zones deviennent plus sèches, et ils restent debout pendant qu’ils marchent entre les arbres jusqu’à ce qu’ils arrivent à la prochaine touffe d’arbres.
Je ne pense pas que nous ayons vraiment un scénario alternatif évolutif très convaincant. Considérez que cette adaptation à la bipédie se déroule sur des millions d’années. Si vous imaginez une créature qui est à quatre pattes, qu’est-ce qui va la faire commencer à marcher debout et le faire assez longtemps pour que le squelette soit modifié par l’évolution pour devenir complètement bipède ? Ils doivent survivre tout au long de ce processus. Très difficile à imaginer.
Des gens comme Darwin ont d’abord spéculé que la bipédie venait de la nécessité d’utiliser des outils ou de transporter des choses, et il est certainement utile de faire ces choses, une fois que vous êtes bipède. Mais qu’est-ce qui va modifier un squelette, modifier la musculature et tout cela, de la même manière que l’évolution nous dit que les primates évoluent au fil des générations?
Ritch-Frel: Prenant ce point quant aux origines de l’apprentissage de la marche, il mène à la discussion sur deux groupes fossiles d’Homo trouvés en Asie du Sud-Est, Homo floresiensis sur l’île de Flores, en Indonésie, et luzonensis dans la grotte de Callao sur l’île de Luçon aux Philippines – et floresiensis avec une hauteur adulte à un peu plus d’un mètre de haut.
Floresiensis a attiré l’attention du public mondial en 2003. On nous a présenté la découverte d’une « créature primitive », une créature qui est plus souvent appelée un « ça » qu’une personne. Les membres les plus curieux du public qui creusent plus profondément dans cette découverte sont généralement informés que ces « hobbits » étaient un produit du nanisme évolutif, souvent trouvé sur les îles, où les grandes créatures sont réduites en taille par des contraintes de ressources et des pools génétiques plus petits. Toujours présent dans les discussions sur floresiensis est l’accent sur leurs petits cerveaux « primitifs ». Nous commençons à apprendre que la taille peut ne pas avoir autant d’importance que la disposition du cerveau lorsque nous nous comparons à nos ancêtres et à leurs capacités fondamentales. (Je vous interrogerai encore à ce sujet plus tard.)
Plus récemment, en 2019, les archéologues ont annoncé une découverte de fossile trouvée à près de 2 000 miles de là, aux Philippines, portant actuellement un nom d’espèce Homo luzonensis qui présente de nombreuses similitudes avec floresiensis.
Jusqu’à leur découverte, on pensait que les premiers hominidés / humains à arriver en Asie du Sud-Est étaient Homo erectus, qui est connu pour avoir quitté l’Afrique il y a environ 2 millions d’années.
Il est à noter que certains experts soutiennent que floresiensis était capable de marcher, mais pas de courir. Et que l’humérus de floresiensis, l’os du bras, était plus long que son fémur, l’os de la jambe supérieure. Ceci est typique d’un type de corps adapté à l’escalade. Les os du poignet indiquent également l’escalade. Ce genre de branche évolutive, je comprends, remonte à quelque part au-delà de 2,5-3 millions d’années, et forcerait à repenser quel style de locomotion des espèces Homo a quitté l’Afrique pour la première fois et peut-être préparer le terrain pour influencer et s’hybrider avec les parents africains qui sont venus après.
Floresiensis / luzonensis est un domaine où il n’y a pas de consensus parmi les experts – et le public pourrait trouver les écoles de pensée illustratives sur les frontières de notre compréhension de l’histoire de l’évolution humaine.
Stringer: Certains experts soutiennent que le scénario le plus convaincant est que le matériau floresiensis est dérivé de l’Homo erectus – qu’il s’agit d’une forme naine d’Homo erectus qui est arrivée à Flores, a subi un nanisme et… a conservé certaines caractéristiques érectes. Nous savons qu’erectus a quitté l’Afrique il y a environ 2 millions d’années. Certaines des caractéristiques dentaires de floresiensis ont été suggérées comme une preuve claire d’une ascendance erectus. Pour que cette idée fonctionne, floresiensis aurait eu besoin d’un ancêtre qui a développé ou redéveloppé indépendamment des caractéristiques basales – des caractéristiques qui ressemblent davantage aux caractéristiques ancestrales d’espèces précédemment développées en Afrique. Comme vous l’avez mentionné, les proportions du corps, le haut du corps qui semble montrer des adaptations pour l’escalade. Peut-être que floresiensis est retourné dans les arbres pour se nourrir. C’est une possibilité.
Ce processus de nanisme aurait dû se produire par la suite dans le processus de migration insulaire en Asie du Sud-Est. C’est un scénario que certaines personnes qui travaillent sur les fossiles d’Homo erectus défendent. C’est une école d’opinion sur floresiensis.
Et d’un autre côté, vous avez des experts qui travaillent dans le sens auquel vous avez fait allusion, et pensent que c’est la preuve d’une sortie pré-erectus de l’Afrique. Un Homo habilis ou même un australopithèque est sorti d’Afrique, a en quelque sorte atteint l’Asie du Sud-Est, en termes de fossiles que nous connaissons, et peut-être aussi à Luçon aux Philippines pour Homo luzonensis. En faveur de cela, nous avons ces caractéristiques basales dans les os du poignet, le bassin et les épaules, et le cerveau plus petit.
C’est un scénario assez convaincant. Mais si vous êtes d’accord avec cela, alors vous devez conclure qu’une évolution convergente, ou indépendamment similaire, de leurs dents vers Homo erectus devait se produire. Les aspects du crâne ressemblent à ceux d’un érectus. Floresiensis a un petit visage qui est niché sous la voûte crânienne, ce qui a nécessité une certaine dérivation. Floresiensis aurait dû avoir à la fois une évolution similaire indépendante à erectus, et un retour à certains éléments plus basaux de leurs ancêtres.
Il existe un point de vue de compromis, selon lequel floresiensis est le produit d’un érectus basal. Certains des fossiles de squelette d’érectus trouvés sur un site appelé Dmanisi dans la Géorgie, ont un cerveau beaucoup plus petit. L’un des fossiles a une taille de cerveau pas trop différente de floresiensis.
Nous pourrions partir d’un érectus plus petit, plus petit cerveau, et peut-être qu’alors il aurait pu arriver à Flores finalement, et a évolué et survécu là-bas pendant plus d’un million d’années. Nous devons garder à l’esprit que nous ne connaissons pas l’anatomie complète de l’érection de toute façon. Alors, à quoi ressemblaient les os du poignet à Dmanisi? Étaient-ils comme ceux trouvés à Flores? Nous ne le savons tout simplement pas encore, car ils n’ont pas été préservés jusqu’à présent.
Dans tous ces cas, vous avez également le mystère sur la manière dont ils sont arrivés à Flores: il n’y a pas de ponts terrestres qui apparaissent lorsque le niveau de la mer baisse pendant les périodes glaciaires. Les gens qui soutiennent que floresiensis était plus étroitement lié aux humains via la ligne erectus suggèrent qu’il y avait une capacité d’utiliser peut-être des embarcations pour se rendre à Flores.
Mais l’autre option est que son arrivée sur Flores était accidentelle. Tectoniquement, cette partie de l’Indonésie est l’une des régions les plus actives au monde, causée par des éruptions volcaniques et des tremblements de terre. Il y a eu un tsunami majeur dans l’océan Indien en 2004. Des personnes ont été retrouvées en mer quelques jours plus tard, survivant sur des touffes de végétation. C’est quelque chose qui s’est produit au cours des 20 dernières années. Lorsque vous avez une échelle de temps de milliers, de centaines de milliers, de millions d’années potentiellement, ces événements « rares » peuvent se produire. Nous savons que c’est le nombre d’autres animaux qui ont dû traverser ces îles entre Java et la Papouasie-Nouvelle-Guinée / Australie.
Il est possible que certains ancêtres de floresiensis se nourrissaient peut-être dans les mangroves de la côte, et qu’un raz-de-marée ait arraché toute une zone, et ils sont laissés là, et d’une manière ou d’une autre miraculeusement quelques semaines plus tard, ils arrivent à Flores ou sur une autre île, car cela aurait pu être accompli par étapes. Il n’est pas nécessaire que ce soit tout droit jusqu’à Flores.
Ritch-Frel: Que le floresiensis ait été abattu à dessein ou par accident, il existe cet autre élément de preuve que nous identifions au progrès humain : la fabrication d’outils en pierre. Les archéologues ont trouvé sur deux sites de l’île de Flores des outils associés à la découpe de viande âgés de 700 000 et même de plus d’un million d’années.
Avec floresiensis, nous avons un corps qui était peut-être incapable de courir, capable de marcher, mais mieux adapté à l’escalade. Nous avons un cerveau décrit comme minuscule, mais capable de fabriquer des outils. En ce qui concerne la découverte en 2013 de l’Homo naledi en Afrique du Sud, nous avons des preuves vieilles de 230 000 à 300 000 ans d’une autre espèce d’Homo qui avait une courbure sur les os des doigts associée aux primates qui passent leur temps à grimper, ainsi qu’une structure osseuse de la main qui permet aux gens d’apporter de la complexité dans leur fabrication d’outils. Il a une structure de pied similaire à la nôtre. Comme floresiensis, naledi a aussi un cerveau beaucoup plus petit que le nôtre, mais aussi comme floresiensis, il a une structure cérébrale similaire. Des outils ont été trouvés dans la région qui, selon les archéologues, pourraient avoir été créés par naledi.
L’équipe archéologique qui travaille sur le site de Naledi nous dit qu’il existe des preuves d’une culture avec des traits que nous et nos espèces cousines reconnaîtrions: retourner dans la même grotte pour déposer leurs morts et utiliser le feu pour y naviguer. Les Néandertaliens ont laissé une trace du dépôt de dizaines de leurs morts dans une grotte en Espagne appelée Sima de los Huesos il y a environ 430 000 ans. Que ce que nous regardons dans ces grottes soient des cas de meurtre de masse ou de rituel ou autre chose, nous n’avons tout simplement pas les preuves pour le dire. Dans la grotte de Bruniquel en France, nous avons des preuves de l’utilisation néandertalienne du feu et potentiellement de l’habitation dans la grotte il y a au moins 175 000 ans.
Se souvenir des morts, bien sûr, n’est pas un phénomène que nous serions les seuls à posséder. Les éléphants visitent et pleurent les restes de leurs proches et des membres du troupeau tout au long du processus de décomposition. Les mères chimpanzés porteront leurs nourrissons morts avec elles pendant des jours.
Stringer : Naledi est très intrigant. Nous pouvons expliquer la survie de floresiensis à long terme et son évolution divergente dans l’isolement, et l’Homo sapiens n’y arrive pas avant peut-être les 50 000 dernières années, puis floresiensis disparaît. Mais dans le cas du naledi, nous l’avons en Afrique du Sud, sur un continent où nous sommes presque sûrs que l’Homo sapiens avait déjà évolué, où d’autres espèces d’Homo, comme le rhodesiensis, étaient présentes. Et pourtant, le naledi survit en Afrique du Sud avec un cerveau de la taille d’un singe avec succès, apparemment, et passe peut-être son temps profondément dans les systèmes de grottes là-bas.
J’ai été l’un des critiques de l’idée de l’élimination intentionnelle de l’enterrement, parce que j’ai soutenu que « dans quelle mesure le comportement d’une créature avec un cerveau de la taille d’un chimpanzé ou d’un gorille pourrait-il être complexe? »
Mais je suis plus qu’heureux d’être surpris par la complexité beaucoup plus grande de l’Homo naledi lorsque sonnt proches d’aboutir des recherches menées par des pairs qui plaident en la faveur de cette complexité (ce qui pourrait être bientôt).
Ritch-Frel: L’accent est mis sur la taille du cerveau de nos proches en public et sur la conversation d’experts sur les origines humaines, pour nous comparer à nos ancêtres et à nos cousins. Dans le cas de floresiensis et naledi, la conversation publique revient sans cesse sur la petite taille de leur cerveau. Naledi avait une taille de cerveau de 600 millilitres; Chacun d’entre nous en a environ 1 300. Serait-ce un peu un leurre en ce qui concerne leurs capacités de base? Devrions-nous mettre davantage l’accent sur la disposition des structures cérébrales centrales? Cela mérite-t-il d’être mis davantage en avant par rapport à nous?
Stringer: Toute la question de la taille du cerveau et de la complexité du comportement, c’est un débat de longue haleine.
Les Néandertaliens et les sapiens ont des cerveaux relativement gros dans la famille des Homo. Vous pouvez voir une corrélation approximative entre la complexité comportementale croissante des outils en pierre et la taille du cerveau. C’est une corrélation approximative, pas une certitude. C’est pourquoi je pense que le naledi va être très important, parce que si l’équipe de recherche démontre la complexité du comportement, je pense que cela mettra certainement un clou dans le cercueil de l’idée qu’un petit cerveau d’hominidé ne peut pas accomplir des choses complexes.
Ritch-Frel: Compte tenu de cela, et en revenant à certaines des morphologies arboricoles conservées, est-il juste de se demander maintenant si l’intelligence que les humains ont tendance à considérer comme leur caractéristique propre et à l’utiliser comme marqueur de notre différence par rapport aux autres animaux a été développée dans les arbres plutôt que exclusivement sur le sol? Nous savons que les jeunes femelles chimpanzés fabriquent des poupées, par exemple, avec lesquelles elles simulent l’éducation des enfants.
Stringer: Je pense que même en regardant les chimpanzés et les gorilles, ils ont une intelligence claire supérieure à celle de la plupart des autres créatures, de la plupart des autres mammifères. C’était certainement là dans l’ancêtre commun. Je pense donc que l’ancêtre commun entre nous et les chimpanzés il y a environ 7 millions d’années avait déjà un comportement complexe et potentiellement même un comportement de fabrication d’outils à ce stade précoce.
Pourquoi pas? Donc je pense que oui, ça aurait pu commencer à se développer dans les arbres. Et comme je l’ai dit, les orangs-outans sont intelligents aussi. Je pense donc que l’ancêtre commun aurait eu ce degré d’intelligence. Mais il y a des arguments selon lesquels, au moment où nous arrivons à l’australopithèque, il y a eu une certaine restructuration du cerveau, ce qui implique peut-être une réorganisation pour une pensée plus complexe.
Ritch-Frel: Nous savons maintenant qu’il y a au moins cinq espèces humaines distinctes qui vivaient sur Terre il y a seulement 70 000 ans: Homo sapiens, neanderthalensis, denisova, floresiensis et luzonensis. Et nous pouvons démontrer à travers plusieurs sources de preuves qu’ils avaient non seulement une anatomie différente, mais qu’ils avaient également des capacités physiques et des traits ou tendances comportementaux variables.
Une espèce humaine de 1 mètre de haut en Indonésie avait un pied qui rendait la course difficile. La recherche nous dit que les Néandertaliens avaient tendance à être agressifs, à être des gens du matin, à souffrir de dépression; qu’ils nous auraient parus dogmatiques, et qu’ils avaient des comportements répétitifs.
En plus de cela, nous savons également que les sapiens à travers la planète portent aujourd’hui du matériel génomique issu de l’hybridation avec au moins six espèces d’Homo, dont certaines que nous pensons avoir disparu en tant qu’espèce indépendante et distincte bien avant 70 000 ans. Deux de ces espèces que nous pouvons nommer, Neandertal et Denisovan, et les quatre autres que les scientifiques n’ont pas encore nommées, mais nous avons des preuves génomiques de ces « ancêtres mystérieux ».
Cela ne fait pas encore partie de la vulgarisation publique, mais pouvez-vous voir un avenir où les gens pourraient s’identifier et identifier leurs comportements comme typiques de leur famille, de leur religion, de leurs origines régionales et aussi de leurs héritages d’espèces ancestrales dans un environnement où la compréhension de nous-mêmes renforce les liens de coopération et nous fournit un cadre universel de relation potentielle?
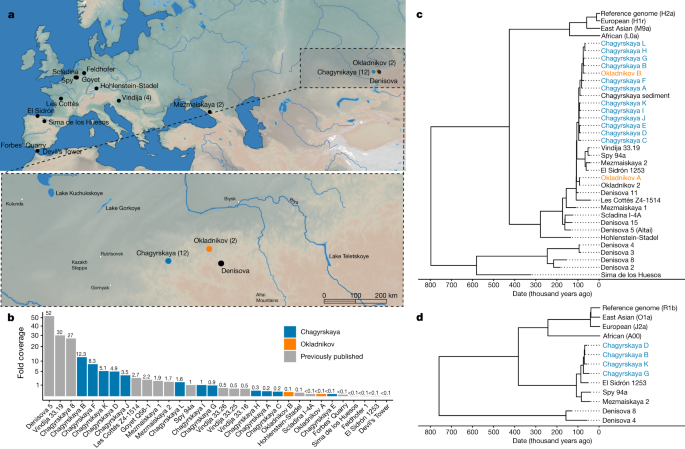
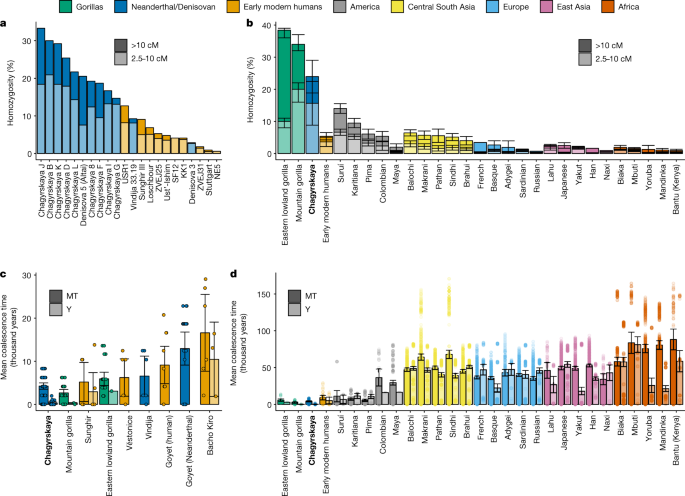
Stringer: Il y a certainement des preuves de métissage sapiens avec les Néandertaliens, et on pense toujours qu’il s’agit d’un groupe assez étroitement lié de Néandertaliens qui s’est hybridé avec Homo sapiens. Mais pour les Denisovans, ce sont au moins trois groupes de population différents de Denisovans qui se sont diversifiés il y a environ 300 000 ans qui se sont croisés avec Homo sapiens dans différentes parties de l’Asie et de l’Asie du Sud-Est.
Et revenons à votre question sur l’identité. Oui, je pense que nous savons par des études ce que l’ADN néandertalien fait en nous aujourd’hui que des morceaux d’ADN néandertalien sont liés, par exemple, au fait que vous soyez une personne du matin ou du soir. Nous savons que certains morceaux d’ADN de Néandertal ont fourni une protection contre COVID. L’âge de la ménopause et le début de la menstruation. Le comportement addictif semble être lié dans certains cas à des morceaux d’ADN de Néandertal.
Il y a des suggestions que l’autisme, la schizophrénie, certainement les maladies auto-immunes, sont également influencés dans une certaine mesure par la présence d’ADN de Néandertal, et probablement nous trouverons des choses similaires pour l’ADN de Denisova. Donc, cela nous affecte certainement, notre biologie de base, nos personnalités.
Et pour les Denisovans, dans certaines populations, il y a deux fois plus d’ADN de Denisova que d’ADN de Néandertal. Les populations d’Asie du Sud-Est ont un ADN néandertalien au même niveau que, disons, les Européens ou les Asiatiques, mais ils ont peut-être 4% d’ADN de Denisova. Donc, théoriquement, nous imaginons que cela va avoir un effet encore plus grand. Nous savons qu’il affecte le système immunitaire, mais il peut aussi avoir d’autres effets.


/image%2F1577277%2F20231227%2Fob_749bcc_grece.jpg)



/image%2F1577277%2F20231029%2Fob_447589_capture.JPG)
/image%2F1577277%2F20231029%2Fob_f0d79a_capture.JPG)
/image%2F1577277%2F20230811%2Fob_a19e3e_capture2.JPG)
/image%2F1577277%2F20230603%2Fob_3d2fdb_capture1.JPG)
/image%2F1577277%2F20230603%2Fob_5b0f4f_capture.JPG)
/image%2F1577277%2F20230603%2Fob_3285a0_capture2.JPG)
/image%2F1577277%2F20230603%2Fob_280253_capture.JPG)
/image%2F1577277%2F20230408%2Fob_d83270_capitalisme-28-1110x611.jpg)
/image%2F1577277%2F20221104%2Fob_2ee625_cadre-3.jpg)

/image%2F1577277%2F20220702%2Fob_530615_safe-image.jpg)
/image%2F1577277%2F20220618%2Fob_32aa8b_129157027-205340437857361-896028546199.jpg)
/image%2F1577277%2F20220618%2Fob_a3ef4c_129336655-205340414524030-575146965581.jpg)
/image%2F1577277%2F20220618%2Fob_deea82_chine-copier.jpg)
/image%2F1577277%2F20220618%2Fob_a9bc9d_chine-onu-11-1024x683.jpg)
/image%2F1577277%2F20220618%2Fob_48772b_chine-onu-14.jpg)
/image%2F1577277%2F20220618%2Fob_7af0dd_chine-puissance.jpg)
/image%2F1577277%2F20220615%2Fob_cd7242_homme-dragon.jpg)
/image%2F1577277%2F20220615%2Fob_4d5a13_homme-dragon2.jpg)